
王晓玲1,2,刘 倩2,韩 伟2,申 琳3,张晓琳2,关 锋1
(1.江南大学 糖化学与生物技术教育部重点实验室 生物工程学院,江苏 无锡 214122;2.国家粮食局科学研究院,北京 100037;3.中国农业大学 食品科学与营养工程学院,北京 100083)
摘 要:棉籽饼粕是一类重要的蛋白饲料资源,但棉粕中有毒物质游离棉酚的存在影响了其在畜牧业中的使用。通过醋酸棉酚与菌株共培养的方法,筛选获得了一株在液体培养条件下棉酚脱除率达90%以上的菌株ST-141,通过16S rDNA序列比对分析和Biolog微生物鉴定,确定该菌株为枯草芽孢杆菌(Bacillus subtilis)。利用该菌株与酿酒酵母N5对棉粕进行混菌固态发酵,30 ℃发酵48 h后棉粕中游离棉酚脱除率达48.5%,同时粗蛋白含量增加19%,氨基酸总量上升22.2%,必需氨基酸含量增加28.4%,表现出良好的棉籽饼粕固态发酵应用潜力。
关键词:棉粕;游离棉酚;枯草芽孢杆菌;固态发酵
我国是产棉大国,2014年全国棉花产量为600多万t,所产棉粕400万t左右。棉粕含有丰富的蛋白质,粗蛋白含量在38%~50%之间,与豆粕相当,此外还富含维生素E和维生素B族中的硫胺素成份,磷含量在1.0%以上[1-2]。但到目前为止,棉粕在饲料中的应用仍存在着添加比例低、适用范围窄等问题,而造成这些问题的最主要原因就是棉粕中有毒物质棉酚的存在。棉酚是一种存在于棉花中的多酚萘衍生物,按其存在形式分为游离棉酚和结合棉酚,一般认为游离棉酚是造成棉粕毒性的主要原因,游离棉酚分子结构中含有活泼的醛基与羟基,能与蛋白等物质发生某些反应,且被动物摄入后不能被完全排出,会在动物体内累积,对动物的生长、发育、繁殖具有一定毒害作用[3-7]。因此世界许多国家和地区对使用棉粕作为饲料时的棉酚含量进行了限定。
目前棉籽饼粕的脱毒方法主要有物理法、化学法和微生物法,物理脱毒法包括热处理法、旋液分离法等,化学脱毒法包括硫酸亚铁法、碱处理法、尿素处理法、氧化法等,这些方法普遍存在着处理后棉粕中某些氨基酸组分损失、后处理困难、能耗高、适口性及色泽较差等问题[8-10]。微生物发酵法脱除棉酚是人们根据牛、羊等反刍动物对棉籽饼粕中的棉酚有较强的耐受能力及霉变的棉籽饼中棉酚含量非常低这些现象的基础上研究发展起来,具有脱毒率高、发酵时间短、成本低、工艺简单等特点,发酵过程还可以改善棉籽饼粕营养及适口性,降解棉粕中不易被动物吸收的大分子蛋白质为活性肽[11-17],如果利用益生菌进行发酵,不仅具有上述优势,对于改善动物肠道微生态也具有一定效果[18]。目前国内利用微生物脱除棉酚的研究主要集中在脱除棉酚菌株的筛选、鉴定以及发酵工艺条件优化等方面,虽已取得了一些成果,仍然存在发酵前后高温处理造成棉粕营养被破坏、微生物脱毒机理不明等问题。
本研究利用醋酸棉酚与菌株96孔板共培养的方法进行高通量筛选,并将筛选获得的菌株与酵母复配,对棉粕进行固态生料发酵,降低游离棉酚含量,改善棉粕营养水平,为发酵棉粕的生产和应用提供新菌株和研究基础。
1.1 材料和试剂
1.1.1 原料和试剂
原料:棉粕,山东青岛,其干物质中游离棉酚含量为1 600 mg/kg,粗蛋白50.2 g/100 g,氨基酸总量41.8 g/100 g;葡萄糖,市售;麸皮,市售。
主要试剂:牛肉膏、马铃薯浸出粉,北京奥博星公司;色谱纯乙腈,Dikma公司;醋酸棉酚标准品,陕西斯诺特生物公司;间苯三酚、PCR产物纯化试剂盒,上海生工生物工程有限公司;Taq DNA聚合酶、dNTP、pMD18-T质粒,TaKaRa公司;引物27F、1495R,英潍捷基公司。
1.1.2 菌株
以本实验室保藏的,从山东、河南采集的棉籽油厂、棉田土壤及棉花植株等样品中分离纯化的菌株,以及中国农业大学食品科学与营养工程学院985平台菌种保藏与应用中心菌株为对象进行筛选。枯草芽孢杆菌ST-141是从中国农业大学食品科学与营养工程学院985平台菌种保藏与应用中心筛选获得的菌株;产朊假丝酵母Y1、酿酒酵母N5、酿酒酵母N31是本实验室筛选保藏的适用于发酵饲料的菌株。
1.1.3 培养基
96孔板筛选液体培养基:牛肉膏1 g/L,蛋白胨10 g/L,氯化钠5 g/L,pH 7.2~7.4,121 ℃灭菌30 min。使用时加入一定量的醋酸棉酚丙酮溶液,使醋酸棉酚终浓度为0.5 g/L。
种子培养基1(培养芽孢杆菌,NB培养基):牛肉膏5 g/L,蛋白胨10 g/L,氯化钠5 g/L,pH 7.2-7.4,121 ℃灭菌30 min。
种子培养基2(培养酵母菌,PDA培养基):马铃薯浸出粉6 g/L,葡萄糖20 g/L,自然pH,115 ℃灭菌30 min。
棉粕固态发酵培养基:棉粕(粉碎过30目筛)95%,葡萄糖2%,麦麸2%,硫酸铵1%。
1.2 实验仪器与设备
高效液相色谱仪:Waters 515/717/474,美国Waters公司;控温控湿摇床:ISF1-X,Adolf Kuhner公司;高速离心机:5810R,德国艾本德股份公司;PCR仪:22331,德国艾本德股份公司;Biolog微生物鉴定仪、Biolog鉴定板:美国Biolog公司;酶标仪:美国BioTek公司;电热恒温培养箱、普通摇床:上海一恒科技公司。
1.3 实验方法
1.3.1 菌株筛选
从菌种平板上挑取单菌落接入5 mL种子培养基1中,37 ℃、200 r/min条件下培养9 h。将加入醋酸棉酚的筛选液体培养基按每孔400 μL分装到96孔深孔板中,然后以5%接种量接入上述种子培养液,37 ℃、250 r/min、85%湿度条件下培养48 h,利用HPLC检测棉酚含量,以未接种并含有相同浓度棉酚的培养基为对照。
1.3.2 棉粕固态发酵
1.3.2.1 固态发酵种子液制备
从菌种平板上挑取单菌落接入5 mL种子培养基1中,37 ℃、200 r/min条件下培养6 h后以1.5%接种量接种到50 mL种子培养基1中,37 ℃、200 r/min条件下培养7 h后作为固态发酵种子液。
1.3.2.2 单菌固态发酵
将活化后种子液以20%接种量接种到棉粕固态发酵培养基中,料水比1∶0.5,pH自然,置于30 ℃恒温培养箱培养48 h,每12 h翻料一次,发酵结束后于45 ℃下烘干6 h粉碎分析测定。
1.3.2.3 混菌固态发酵
同单菌发酵条件,两种或三种不同菌株按等比例接种。
1.3.3 棉酚的提取和检测
1.3.3.1 液体培养醋酸棉酚的提取和检测
提取:取待测样品,14 000 r/min条件下离心5 min,弃上清,加入等体积丙酮,充分混合使其完全浸提。14 000 r/min条件下离心5 min,取其上清进行HPLC检测,对照做同样处理。
检测:安捷伦C18柱(5.0 μm,150×4.6 mm);流动相,乙腈∶水=80∶20,磷酸调至pH 3;流速1.0 mL/min;进样量10 μL;检测波长235 nm。
1.3.3.2 棉粕中游离棉酚的提取和检测
游离棉酚提取:美国油脂化学家协会明确定义“凡能被70%丙酮水溶液提取的棉酚及其衍生物统称为游离棉酚”,因此选用70%丙酮水溶液冰水浴超声90 min提取游离棉酚,料液比为1∶15。
检测:采用间苯三酚法[19]。
1.3.4 菌种鉴定
1.3.4.1 菌株16S rDNA的PCR扩增和序列分析
PCR采用细菌16S rDNA通用引物27F(5′-GAGAGTTTGATCCTGGCTCAG-3′)和1495R(5′-CTACGGCTACCTTGTTACGA-3′)。利用Promega公司的基因组DNA纯化试剂盒提取所筛选菌株的基因组DNA,以基因组DNA为模板进行PCR,PCR扩增条件:95 ℃预变性5 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1.5 min,30个循环,72 ℃延伸5 min。PCR扩增片段纯化后连接到pMD18-T载体上,转化E.coli DH5α。重组质粒由英潍捷基(Invitrogen)公司进行测序,所得序列分别于NCBI和Ez BioCloud网站进行核苷酸比对分析。
1.3.4.2 Biolog菌株鉴定
采用Biolog GENⅢ微生物鉴定板,根据Biolog使用手册中步骤进行操作,分别在4~6 h、16~24 h读板。
1.3.5 生长曲线测定
将活化好的菌株接种到NB培养基中,使接种后培养基OD600=0.05(用未接种的NB培养基做空白对照),于37 ℃、200 r/min振荡培养,每隔1 h取样,600 nm波长(对细胞密度大的培养液适当稀释后测定)进行光电比浊测定,同时进行菌落计数。
1.3.6 平板菌落计数
菌落总数的测定采用GB 4789.2—2010。
1.3.7 棉粕中营养物质的测定
粗蛋白测定采用GB/T 6432—1994;18种氨基酸测定采用GB/T 18246—2000。
2.1 棉酚脱毒菌株筛选及菌种鉴定
利用96孔板棉酚与菌株共培养的方法,筛选具有棉酚脱除能力的菌株,通过HPLC检测,发现一株棉酚脱除能力较高的菌株ST-141,该菌株液体培养48 h后培养基中的棉酚含量与对照相比下降了92%(图1)。
a 发酵前HPLC检测图; b 发酵后HPLC检测图
图1 菌株ST-141与醋酸棉酚共培养前后HPLC检测图
对菌株ST-141进行菌种鉴定,利用细菌通用引物扩增获得了该菌株的16S rDNA核苷酸片段,大小为1 516 bp,通过BLAST核苷酸比对分析表明其与一株枯草芽孢杆菌的16S rDNA序列同源性为99%(Genbank accession no.AB480760.1);Ez Taxon比对发现其与Bacillus subtilis subsp.subtilis NCIB 3610(ABQL01000001)16S rDNA序列相似度为99.8%。利用MEGA 5.0构建16S rDNA进化树,发现ST-141与Bacillus subtilis位于同一分支,说明两者亲缘关系较近(图2)。

图2 基于16S rDNA 基因序列采用临近法构建的
ST-141和相近菌株的系统发育进化树
利用Biolog GENⅢ微生物鉴定板对该菌株进行进一步鉴定。Biolog自动微生物鉴定系统是基于95种碳源或化学敏感物质的利用为原理进行菌种鉴定,再利用独创的算法模型,用可能性(probalitity,PROB)、相似性(simirity,SIM)和位距(distance,DIST)三个参数判定鉴定结果,其中SIM值是判断结果是否可信的重要参数。Biolog系统规定,细菌培养4~6 h,其SIM≥0.75,或者培养16~24 h时,SIM≥0.5时,系统自动给出鉴定结果为种名,当SIM值小于0.5,但鉴定结果中属名相同的结果的SIM值之和大于0.5时,自动给出的鉴定结果为属名。将菌株ST-141培养17 h时检测,Biolog系统显示SIM为0.59,PROB为80%,DIST为3.624,鉴定结果为枯草芽孢杆菌,由于此时SIM值大于0.5,因此该种名可信。
综合分子鉴定和Biolog鉴定结果,可确定ST-141为一株枯草芽孢杆菌。枯草芽孢杆菌繁殖迅速,是一种益生菌,属于我国《饲料添加剂品种目录(2013)》内可允许添加微生物,具有抑制有害微生物生长、调节动物肠道微生态平衡等优点,在饲料、制药、污水处理等方面有广泛应用。
2.2 菌株ST-141的生长曲线
为了解枯草芽孢杆菌ST-141的生长周期,为后续棉粕发酵实验提供依据,绘制菌株ST-141的生长曲线并进行活菌计数。在NB培养基中,37 ℃、220 r/min条件下培养,不同时间取样分别测定OD600值并进行活菌计数,绘制生长曲线。从图3生长曲线可以看出,0~2 h时菌株处于延滞期,2~14 h为对数生长期,随着时间延长,OD值也不断增高,14~16 h菌株生长进入稳定期,之后OD值开始下降,进入衰亡期。而根据活菌计数结果,2~7 h活菌数处于对数增长期,7 h后进入稳定期,此时活菌数最高,达到6.1×108CFU/mL,因此在后续的固态发酵中种子液培养时间选择为7 h。

图3 枯草芽孢杆菌ST-141的生长曲线
2.3 棉粕固态发酵
利用枯草芽孢杆菌ST-141对棉粕进行固态生料发酵,结果表明经过48 h的发酵后棉粕中游离棉酚的含量降为950 mg/kg,与发酵前相比下降了37%,说明该菌株在棉粕固态发酵时具有一定的棉酚脱除能力。在以往的文献中,一些研究者报道固态发酵时棉酚脱除率可达90%以上[20-21],主要是由于这些研究中往往采用了高温高压灭菌熟料发酵或发酵后高温烘干的方式,通过对棉粕进行高温处理降低了棉酚的含量,而真正微生物发酵的作用较小。本研究采用生料发酵,减少了操作工序,降低能耗,同时也减少高温所造成的营养物质损失,具有一定优势。
目前微生物发酵生产蛋白饲料的方式已从单菌发酵趋向于复合菌株协同发酵,利用不同微生物之间的协同性和互补性,从总体上发挥正组合效应[22-24]。利用ST-141菌株对棉粕进行固态发酵,虽然能在一定程度上降低棉酚含量,但仍存在棉酚含量偏高、发酵后有异味等问题。为改善棉粕风味,进一步降低棉酚含量,增加棉粕中益生菌含量,提高其饲用品质,选用实验室保藏的产朊假丝酵母Y1、酿酒酵母N5、酿酒酵母N31与ST-141进行固态混菌发酵(图4)。结果表明固态混菌发酵后游离棉酚的脱除率与单菌发酵相比有不同幅度的提高,其中ST-141与酿酒酵母N5协同发酵时棉酚脱除率最高,为48.5%,比ST-141单菌发酵时棉酚脱除率提高了11.5个百分点,同时发酵后棉粕气味酸香,与枯草芽孢杆菌单独发酵相比有所改善。

图4 ST-141与不同酵母菌协同发酵时的棉酚脱除率
2.4 发酵棉粕粗蛋白及氨基酸含量变化
以枯草芽孢杆菌ST-141与酿酒酵母N5作为实验菌株进行棉粕固态发酵,对发酵前后粗蛋白及氨基酸含量进行检测。发酵后棉粕中粗蛋白含量为59.8 g/100 g,与发酵前50.2 g/100 g相比提高19%;氨基酸含量变化如图5所示,发酵后棉粕与发酵前相比,18种氨基酸含量皆有提高,氨基酸总量由40.8 g/100 g上升到51.1 g/100 g,提高22.2%,8种必需氨基酸总量由12.4 g/100 g增加到16.6 g/100 g,提高28.4%。利用枯草芽孢杆菌和酿酒酵母对棉粕进行固态发酵,不仅降低了游离棉酚含量,而且提高了棉粕中粗蛋白和氨基酸的含量,改善了棉粕的营养水平。
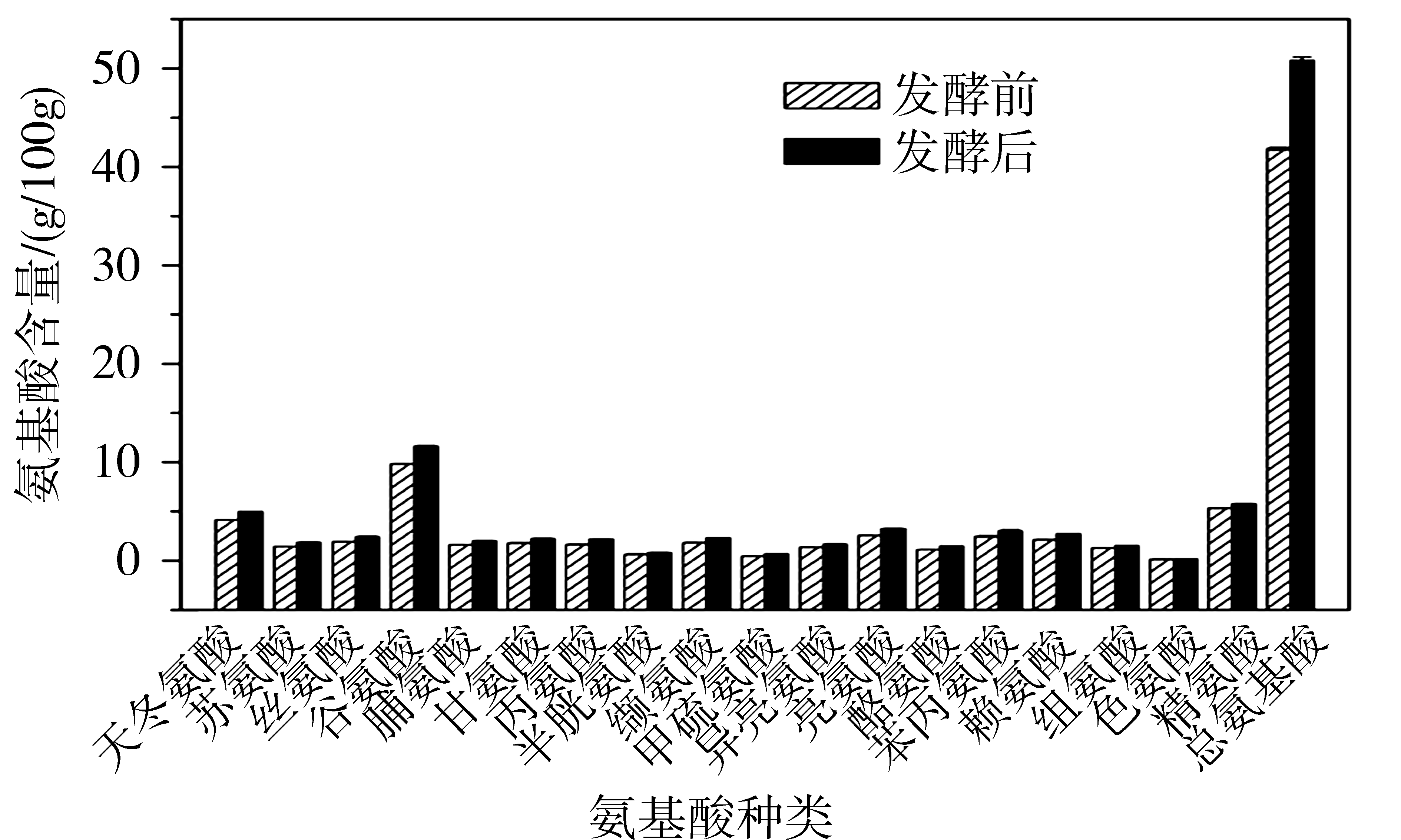
图5 棉粕固态发酵前后氨基酸含量变化
本研究利用菌株与醋酸棉酚共培养的方法筛选具有棉酚脱除能力的菌株,获得一株在液体培养条件下棉酚脱除率可达到92%的菌株ST-141,通过16S rDNA序列比对和Biolog自动微生物鉴定系统分析,将该菌株鉴定为枯草芽孢杆菌。利用ST-141与一株酿酒酵母进行棉粕固态混菌发酵,48 h后棉粕中游离棉酚脱除率为48.5%,同时粗蛋白含量增加19%,氨基酸总量上升22.2%。本研究采用生料发酵,并且发酵后45 ℃烘干,避免了高温对棉粕营养物质的破坏,通过微生物的作用降低游离棉酚含量,减少其毒性,同时对棉粕的营养水平和适口性有一定改善作用,为棉粕的固态发酵提供了新菌株和研究基础。
参考文献:
[1]刘靖,张石蕊.蛋白质饲料资源的合理利用及开发对策[J].中国畜牧兽医文摘,2009,30(5):43-46.
[2]中国饲料数据库.中国饲料成分及营养价值表(第25版)[J].中国饲料,2014,(21).
[3]Tang J W,Sun H,Yao X H,et al.Effects of Replacement of Soybean Meal by Fermented Cottonseed Meal on Growth Performance,Serum Biochemical Parameters and Immune Function of Yellow-feathered Broilers[J].Asian Australasian Journal of Animal Sciences,2012,25(3):393-400.
[4]王飞,米海峰.棉籽饼粕中棉酚对动物肌体的毒害作用及其预防
措施[J].粮食与饲料工业,2003(8):27-28.
[5]Randel R D,Chase C C,Wyse S J.Effects of Gossypol and cottonseed products on reproduction of mammals[J].J Anim Sci,1992,70(5):1628-1638.
[6]Fombad R B,Bryant M J.An Evaluation of the Use of Cottonseed Cake in the Diet of Growing Pigs[J].Tropical Animal Health & Production,2004,36(3):295-305.
[7]候红利,罗宇良.棉酚毒性研究的回顾[J].水利渔业,2005,25(6):100-102.
[8]赵顺红,张文举.棉籽饼粕生物脱毒的研究进展[J].畜牧与饲料科学,2007,28(1):54-55.
[9]吕慧.棉籽饼粕的脱毒方法的比较[J].中国棉花加工,2009,1(1):29-30.
[10]于涛,申德超,庞芳.棉籽粕脱毒方法的研究进展[J].山东理工大学学报:自然科学版,2006,4(4):88-92.
[11]Yang X,Guo J,Sun J.Biodegradation of free-gossypol by a new fungus isolated from cotton planted soil[J].African Journal of Microbiology Research,2011,5(19):3066-3072.
[12]Yang X,Sun J,Guo J,et al.Identification and proteomic analysis of a novel gossypol-degrading fungal strain[J].Journal of the Science of Food & Agriculture,2012,92(4):943-951.
[13]Meleigy S A.Reduction of free gossypol levels in cottonseed meal by microbial treatment[J].International Journal of Agriculture & Biology,2008,10(2):185-190.
[14]金红春,兰时乐,胡毅等.棉粕发酵前后营养成分变化研究[J].饲料工业,2011,32(13):19-23.
[15]邓露芳,范学珊,王加启.微生物发酵粕类蛋白质饲料的研究进展[J].中国畜牧兽医,2011,38(6):25-30.
[16]李旋亮,吴长德,李建涛,等.微生物发酵饲料的研究与应用[J].饲料博览,2010,2(2):27-29.
[17]王子强.微生物发酵饲料研究进展[J].畜牧与饲料科学,2010,31(4):34-36.
[18]李永凯,毛胜勇,朱伟云.益生菌发酵饲料研究及应用现状[J].畜牧与兽医,2009,41(3):90-93.
[19]郑清梅,温茹淑,韩春艳.间苯三酚法快速测定棉粕中游离棉酚的方法研究[J].饲料工业,2010,31(3):42-44.
[20]魏二虹,张文举,刘东军,等.复合发酵对棉籽饼混合底物中FG和BG含量的影响[J].饲料工业,2011,32(20):34-38.
[21]朱德伟,刘志鹏,蔡国林,等.高效降解棉酚菌种的筛选及棉粕发酵脱毒工艺研究[J].中国油脂,2010,35(2):24-28.
[22]邱良伟,李爱科,程茂基,等.两种不同发酵工艺的棉籽粕营养价值的研究[J].饲料工业,2012,33(13):32-36.
[23]张庆华,赵新海,钟丽娟,等.三菌株协同固态发酵对棉粕脱毒效果及其生物活性的影响[J].饲料工业,2007,28(18):37-38.
[24]诸葛斌,刘俊,方慧英.混菌发酵改良棉粕蛋白工艺及协同作用研究[J].中国生物工程杂志,2011,31(9):62-68.●![]()
WANG Xiao-ling1,2,LIU Qian2,HAN Wei2,SHEN Lin3,ZHANG Xiao-lin2,GUAN Feng1
(1.Key Laboratory of Carbohydrate Chemistry & Biotechnology Ministry of Education,School of Biotechnology,Jiangnan University,Wuxi,Jiangsu 214122;2.Academy of State Administration of Grain,Beijing 100037;3.College of Food Science & Nutritional Engineering,China Agricultural University,Beijing 100083)
Abstract:Cottonseed meal is an important protein feed resource,however the existence of free gossypol affects its application in animal husbandry.In this paper,a strain ST-141,which could remove more than 90% of free gossypol in the liquid fermentation was isolated and identified asBacillus subtilis through 16S rDNA sequence analysis and Biolog microbial identification.Then the ST-141,mixed with the strain of Saccharomyces cerevisiae N5,were used for the solid state fermentation of cottonseed meal.After incubated at 30 ℃ for 48 h,the removal rate of free gossypol reached 48.5%,and the contents of crude protein,total amino acid and essential amino acid were also increased by 19%,22.2% and 28.4% respectively,showing high application potential.
Key words:Cottonseed meal;Free gossypol;Bacillus subtilis;Solid state fermentation
收稿日期:2015-06-26
基金项目:中央级公益性科研院所基本科研业务费专项资金课题(ZX1505);“十二五”国家科技支撑计划项目(2011BAD 26B01-3)
作者简介:王晓玲,1991年出生,女,硕士研究生.
通讯作者:关锋,1978年出生,男,教授.
中图分类号:TS 201.3
文献标识码:A
文章编号:1007-7561(2016)01-0081-05